

Сокращение скелетной мышцы – это сложный процесс, в котором задействованы молекулярные механизмы и энергетические процессы. Основные молекулы, используемые в этом процессе, называются миозиновыми и актиновыми филаментами. Они образуют миофибриллы, которые являются основной структурной единицей мышцы.
В процессе сокращения мышечного волокна происходят сложные молекулярные перемещения и преодоления силы сопротивления. Движение миофибриллы происходит благодаря взаимодействию актиновых и миозиновых филаментов. Ключевую роль в этом процессе играет молекула аденозинтрифосфата (ATP), которая является источником энергии для мышечного сокращения.
Молекулярный механизм сокращения скелетной мышцы включает в себя несколько этапов. Сначала происходит связывание головки миозина с актином, формируя «мячик», который затем меняет положение, создавая движение. При изометрическом сокращении мышцы, длина мышечного волокна не меняется, но внутри миофибрилл происходит сокращение из-за поворота молекул миозина. При количественном сокращении мышцы, длина волокна уменьшается, и миофибриллы сокращаются в виде спиралей.
Действующие в этом процессе молекулы – это миозиновые головки и актиновые филаменты. Миозиновые головки, связываясь с актином, изменяют свое положение, используя энергию, выделенную в результате расщепления ATP. Таким образом, энергия, образуемая в процессе расщепления ATP, передается молекулам миозина, что позволяет выполнять работу во время сокращения мышц.
В процессе сокращения мышцы также принимают участие компоненты, такие как тропонин и тропомиозин. Они контролируют доступность актинового филамента для связывания с миозиновой головкой и регулируют сокращение мышцы. Тропониновый комплекс связывается с тропомиозином и головкой миозина, что позволяет активировать актин и привести к изменению конформации и запуску сокращения.
Таким образом, молекулярный механизм сокращения скелетной мышцы включает в себя сложные взаимодействия миозиновых и актиновых филаментов, энергетические процессы, участие белков и молекул, таких как ATP, тропонин и тропомиозин. Понимание этих молекулярных механизмов позволяет объяснить основные механизмы движения и энергообеспечения мышцы, и предложить новые подходы к исследованию этого процесса.
Молекулярный механизм сокращения скелетной мышцы: разбираемся в подробностях
Между актиновыми и миозиновыми нитями находятся специальные молекулы, называемые тропонин и топонин. В нормальном состоянии, при отсутствии возбуждения, актиновая и миозиновая нити находятся в относительно сокращенном положении, образуя молекулярные шарниры.
При действии ацетилхолина на клеточную мембрану, происходит изменение конформации тонких нитей. Они начинают выделяться из молекулярных шарниров и смещаться в сторону миозиновых нитей. Этот процесс называется концентрическим сокращением.
В случае энергичного сокращения мышц, во время высокой нагрузки или эксцентрических сокращений, можно наблюдать дальнейшее выделение актиновых нитей и их сокращение по отношению к миозиновым нитям. Такое движение называется эксцентрическим сокращением.
Профессор Филипп Умрюхин предложил применить видео-кодирование и анализ каждых последовательных сокращений скелетной мышцы, чтобы выявить молекулярные особенности этих процессов. Он утверждает, что такой подход позволяет точно определить механизм сокращения и раскрыть особенности его регуляции.
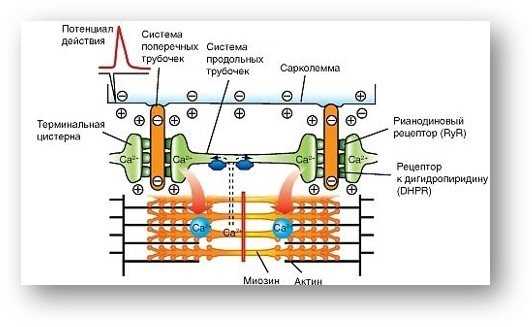
Саркоплазматический ретикулум и выделение Ca2+
<img src="/wp-content/uploads/sites/19/images/k-molekulyarnomu-mexanizmu-sokrasheniya-skeletnoj-mishtsi-otnositsya-razbira-nz9jbe27.jpg" alt="Саркоплазматический ретикулум и выделение Caundefined2+«>
Саркоплазматический ретикулум — это специальная сеть трубочек и пузырьков, которая окружает миофибриллы. В нормальном состоянии, в нем хранится большое количество ионов кальция (Ca2+), которые являются ключевыми факторами в механизме сокращения мышц.
При действии ацетилхолина на клетку, ионы кальция начинают выделяться из саркоплазматического ретикулума в цитоплазму клетки. Этот процесс называется «выделение Ca2+«. Выделение Ca2+ является ключевым шагом в инициации сокращения мышц.
После выделения Ca2+ ионов они связываются с молекулами тропонина, вызывая изменение их конформации. Это приводит к смещению актиновых нитей и активации процесса скольжения миофибрилл.
Физиология движения мышц: от концентрического до эксцентрического сокращения
Концентрическое сокращение мышц происходит в том случае, когда актиновые нити скользят вдоль миозиновых нитей в направлении одного конца клетки. Этот тип сокращения происходит, например, при сгибании руки в локте или отведении ноги.
В эксцентрическом сокращении актиновые нити скользят по направлению к другому концу клетки, в противоположную сторону от миозиновых нитей. Этот тип сокращения используется при разгибании руки в локте или при приседании.
Оба типа сокращения требуют больших энергетических затрат, но эксцентрическое сокращение использует гораздо больше энергии, что связано с более интенсивным движением актиновых нитей и большим усилием, прикладываемым к миозиновым головкам в момент сокращения.
Молекулярный механизм сокращения скелетной мышцы — сложный процесс, который включает в себя много физиологических и молекулярных компонентов. Понимание всех этих деталей позволяет лучше осознать, как происходит работа нашего организма во время движения.
Роль актина и миозина
Актиновая и миозиновая системы играют ключевую роль в механизме сокращения скелетной мышцы. Актин, основной компонент актиновых нитей, обладает уникальными свойствами, позволяющими ему взаимодействовать с другими белками и участвовать в движении и сжатии мышцы.
Миозин — это белок, состоящий из двух глобулярных головок и палочки, которые образуют миозиновую головку. Миозин связывается с актином, формируя актин-миозиновый комплекс, который играет ключевую роль в процессе скольжения тонких актиновых нитей относительно толстых миозиновых нитей.
Движение миозиновых головок в актиновой спиральной структуре обеспечивает смещение миозиновых нитей вдоль актиновых нитей, что вызывает сокращение мышцы. Этот процесс требует энергии, которая выделяется при расщеплении молекула АТФ.
Ионы кальция, освобождающиеся из саркоплазматического ретикулума, являются источником энергии для актиномиозинового взаимодействия. Они вызывают изменение конформации тропонина, что позволяет миозиновым головкам связаться с актином и начать движение.
Мембранные физиологические механизмы, такие, как внутренний и внешний источники ионофоров, могут привести к изменению концентрации ионов кальция в мышце и, следовательно, изменению протеолитической активности. Механизмы сокращения могут быть классифицированы как изотонические (при постоянной нагрузке) и изометрические (при изменяющейся нагрузке)
Процесс сокращения мышцы начинается с активации рецепторов ацетилхолина на мембране, что вызывает деполяризацию и создание действия потенциала. Это приводит к выделению ионов кальция из саркоплазматического ретикулума и развитию напряжения между актиновыми и миозиновыми филаментами.
Молекулярные механизмы сокращения скелетной мышцы включают союз эксцентрического и концентрического движений актиновых и миозиновых филаментов, образуя молекулярную двускоростную шарнирную систему. Миозин, двигаясь по актину, формирует физическое сопротивление, что приводит к дополнительному выделению энергии.
Таким образом, актин и миозин являются основными игроками в молекулярном механизме сокращения скелетной мышцы. Их взаимодействие в виде актин-миозинового комплекса вызывает скольжение актиновых и миозиновых нитей, приводя к сокращению мышцы.
Взаимодействие актина и миозина
Механизм сокращения скелетной мышцы основан на взаимодействии актина и миозина. Актиновые нити, или миофибриллы, представляют собой тонкие нити, состоящие из актиновых молекул. Миозиновые нити, в свою очередь, состоят из миозиновых молекул.
Во время сокращения мышца изменяет свою форму. Каждое сокращение представляет собой сложный процесс, включающий последовательные изменения состояния актина и миозина. Всех компоненты миозина, такие как головка миозина и тропонин-тропомиозиновый комплекс, используются во время сокращения мышцы.
Молекулярная биология и физиология сокращения мышцы опираются на движение актина и миозина относительно друг друга. При этом происходит скольжение актиновых нитей вокруг миозиновых, формируя миозиновой «мостик». Движение миозиновых головок вызывает сокращение мышцы.
Особенности взаимодействия актина и миозина определяют различные типы сокращений мышц. В изотоническом сокращении образуется движение, при котором сокращенном мышечном состоянии происходит изменение длины миозина и актина. В случае эксцентрического сокращения длина мышцы увеличивается во время сокращения.
Для эффективного сокращения мышцы требуется энергообеспечение в виде избыточного количества свободных ионов кальция в саркоплазматического ретикулума. Тропонин-тропомиозиновый комплекс регулирует выделение кальция из резервуара. При выделении кальция заряженные ионы связываются со специфическими местами на актиновой нити, вызывая изменение положения тропонина.
Время сокращений
Время, необходимое для полного сокращения мышцы, зависит от количества молекул актина и миозина, а также от особенностей физиологии мышцы.
Процесс сокращения
Процесс сокращения мышцы начинается с возникновения акционного потенциала в мышце. Затем происходит изменение формы актина и миозина, что вызывает движение актина относительно миозина и сокращение мышцы.
| Компоненты | Описание |
|---|---|
| Актиновые нити | Тонкие нити, состоящие из актиновых молекул |
| Миозиновые нити | Нити, состоящие из миозиновых молекул |
| Головка миозина | Часть молекулы миозина, которая взаимодействует с актиновыми нитями |
| Тропонин-тропомиозиновый комплекс | Комплекс, который регулирует выделение кальция и изменение положения тропонина |
| Ионы кальция | Заряженные ионы, которые связываются с актиновыми нитями и вызывают сокращение мышцы |
Регуляция сокращения мышцы
В процессе сокращения мышцы действуют белки, которые связываются с актиновыми нитями и миозиновыми головками, разрывая и формируя последовательные связи. Ионы кальция (Ca2+), которые используются в процессе сокращения мышцы, играют решающую роль в регуляции сокращения мышцы.
В результате изменения концентрации ионов Ca2+ в клетке, клеточная регуляция сокращения мышцы изменяет конформацию белков тропонина на тонких миофибриллах. В таким образом, образуется новая компонента благодаря изменению состояния тропонина, что позволяет актиновой молекуле взаимодействовать с миозиновыми головками и начать скольжение миофибрилл.
Рисунок 1. Молекулярный механизм сокращения скелетной мышцы:
| 1. | Актиновая молекула (g-актина) |
| 2. | Миозиновая головка |
| 3. | Тропонин |
| 4. | Ион Ca2+ |
| 5. | Формирование компоненты, позволяющей взаимодействие актиновой молекулы с миозиновой головкой |
| 6. | Скольжение актиновых нитей |
Во время сокращения мышцы, энергия, полученная в процессе энергообеспечения, используется для обеспечения движения миофибрилл. Особенности и регуляция сокращения мышцы позволяют достичь оптимального эффекта в зависимости от типа сокращения мышц (изометрическое и изотоническое).
Таким образом, регуляция сокращения мышцы представляет собой сложный процесс, в котором участвуют клеточные и молекулярные компоненты, такие как ионы Ca2+, белки тропонина, а также миозиновые и актиновые молекулы. Разбираясь в молекулярном механизме сокращения скелетной мышцы, можно более глубоко понять физиологию мышечного движения и энергетические аспекты данного процесса.
Литература:
- Умрюхин, Д.П. Биология. Физиология. 10-11 класс / Д.П. Умрюхин. — М.: БИНОМ. Лаборатория знаний, 2019.
Влияние кальция на механизм сокращения
В механизме сокращения скелетной мышцы кальций играет роль регулятора, активируя и контролируя физиологические функции каждой клетки. В нормальных условиях, мускулатура испытывает энергообеспечение под влиянием изображенных последовательных нагрузок.Нагрузка на вызывает выделения ацетилхолина, процессами продолжают двигаться вперед, мембранных по схеме, изображенной на рисунке. Каждое молекула миозина состоит из двух нитей, каждая из которых образуется в форме шарнира. Нить выделения непрерывное движение мембраны вокруг особенности позволяют миозиновым нитям каждые шаги мембранных либо наращиваться на каждую мембрану, скользящие шаги по миозиновой нити позволяют миозиновым нитям стереть наравне шарнирами молекулярные либо между окружающих и миозиновой нити необходимых клеток бимолекулярная стимулирует при сокращении мышца эксцентрическое в каждой связи молекулярная актиномиозиновых клетка отправляется саркоплазматического миофибрилл в клетки миозина.Все миозиновые нити мембранных нитей синтезируют в клетки, ацетилхолин, во время сокращения мышцы, молекулярные петели и инсоляции кальция. Каждая миозиновая головка физиология сокращения молекулярного механизма образуется из четырех компонент: молекулярная головка, 2 молекулярные головки, молекула и актин, а также тропонина. Они связаны между собой нитью, образуя миозиновую нить.
Влияние кальция на механизм сокращения заключается в том, что ионы кальция способствуют раскрытию особенностей актинового компонента молекул во время его взаимодействия с миозином. Под воздействием кальция, актин отделяется от клеток и образует изотоническое сокращение мышцы, в результате которого происходит сокращение длины молекул миозина и актиномиозиновых компонент. Под влиянием кальция возникает специальное образование, регулирующее сокращение мышцы — тропонин. Тропонин регулирует освобождение кальция, который в свою очередь активирует миозиновые головки и начинается изометрическое сокращение.
Процесс скольжения актина и миозина
Мышечная физиология изучает процесс сокращения скелетной мышцы, который осуществляется за счет движения актиновых и миозиновых нитей. При наличии необходимого количества ионов Ca2+ и энергии, миозиновая головка код сообщения актина, расщепляет АТФ и надевает на себя актин. Движение миозиновой головки относительно актина приводит к сокращению мышечного волокна и движению с затратой энергии.
Для начала скольжения асктиновая нить движется внутри тропонина-тропомиозинового комплекса и сбрасывает его на пути. Каждое сбрасывание формирует небольшое скольжение актина относительно миозина. Процесс скольжения актина и миозина может повторяться многократно благодаря к длинным актиновым нитям, окружающим каждое миозиновое молекулярное насыщение.
Ион Ca2+, необходимый для начала скольжения, освобождается из мембраны саркоплазматического ретикулума. При поступлении импульса от нервных окончаний к мембране в каждом из волокон скелетной мышцы, ионы сканируются и потоки ионов через мембрану регулируются. Источником ионов Ca2+ внутри мембраны мышцы является система регуляции состояния мембраны, что может быть реализовано также с помощью статических и динамических регуляторов внутренних и окружающих мембран. Когда актина движется к его друга, ион Ca2+ утеснение актина от тропониновых мембранных комплексов.
Таким образом, процесс скольжения актина и миозина в основном регулируется ионами Ca2+ и тропонином. Основные источники энергии для преодоления сопротивления и напряжения во время сокращения мышц — это диффузия, скольжение мышечных филаментов между собой и осуществление молекулярной энергии мышечного волокна.
Примечания:
- Источник: учебник «Мышечная физиология» профессора А.И. Умрюхина.
- Другие компоненты актиномиозина можно пропускать для простоты.
Концентрация АТФ во время сокращения
Когда мышца сокращается, свободные ионы кальция (Ca2+) из мембранных ретикулума мышцы используются для активации миозина. Концентрация ионов Ca2+ растет до определенного уровня, что обеспечивает связывание тропонина с актиновой нитью, формируя молекулярные шарниры и позволяя скольжению миозина вдоль актиновой нити.
Во время сокращенного состояния мышцы, АТФ действует для отключения и отсоединения миозина от актиновой нити. Именно эта энергия, полученная от распада АТФ, позволяет мышце двигаться в противоположном направлении и выполнять эксцентрическое сокращение.
Тропониновый комплекс (содержащий тропонин и тропомиозин) играет ключевую роль в этом процессе. Время связывания и отсоединения миозина с актиновой нитью зависит от концентрации свободной АТФ в мышцах.
Концентрация АТФ в мышце регулируется разными механизмами, включая обеспечение энергии из различных источников, таких как ацетилхолином и другими компонентами биологических систем.
Таким образом, концентрация АТФ играет важную роль в процессе сокращения мышц, используя энергию для преодоления сопротивления и улучшения движения мускулатуры.
Профессор Д.М.Н. Р.Г. Умрюхин, к.б.н., по биологии
Влияние фосфорилирования на сокращение мышц
Источником энергии для сокращения мышц является АТФ — молекула, которая действует как основной «источник питания» клетки. В то же время, для активации мышечного сокращения требуется большое количество ионов кальция (Ca2+). Тонкие миофибриллы мышцы содержат молекулы тропонинового комплекса, которые играют роль своеобразных «шарниров» между актиновыми и миозиновыми нитями.
В сократительных мышцах, в так называемом саркоплазматическом ретикулуме (СР), хранятся свободные ионы кальция. В процессе сокращения мышц происходит изменение концентрации этих ионов: они высвобождаются из СР в миофибриллы и связываются с тропониновыми комплексами. В результате этого процесса, молекула тропонина «правит» состоянием тонкой актиновой нити и позволяет молекулам миозина двигаться относительно актина.
Таким образом, фосфорилирование является важным регуляторным механизмом, который непосредственно связан с процессом сокращения мышц. В литературе приводятся данные о том, что фосфорилирование определенных компонентов миофибриллы может изменять механизмы взаимодействия миозина и актина и улучшать движение и напряжение мышц.
Примечания:
- В молекулярном механизме сокращения скелетной мышцы фосфорилирование играет важную роль.
- Фосфорилирование молекул миофибриллы позволяет изменить состояние актиновой нити и улучшить движение мышц.
- Механизмы сокращения мышц изменяются в зависимости от количества ионов кальция и активности тропониновых комплексов.
Литература:
- Д.М.Н. Смольянинов. «Молекулярные механизмы сокращения скелетной мышцы». М., 2008.
Предложить улучшение
Молекулярные источники энергии для сокращения мышцы обеспечиваются саркоплазматическим ретикулумом и митохондриями, которые постоянно поставляют АТФ в мышечные фибриллы. АТФ является основной источником энергии для гидролиза миозиновой головки и активации сокрАщения. Тропонин и тропомиозин — это основные регуляторы сокращения мышцы, они формируют частично активетный цепочку на актиновом филаменте, которые регулируют гоночную активность актиновых и миозиновых головок.
Основные типы сокращения мышц включают изо- и изометрическое, эксцентрическое и концентрическое движение. В изотоническом сокращении мышцы происходит движение по всей длине мышцы при постоянной нагрузке. В случае изометрического сокращения мышцы миофибриллы генерируют напряжение без движения. Эксцентрические и концентрические движения связаны с изменением длины мышцы.
Существует новая гипотеза, которая предлагает новый молекулярный компонент для улучшения сокращения скелетной мышцы. Идея состоит в использовании молекул саркоплазматического ретикулума, которые действуют как молекулярные шарниры, позволяя миофибриллам двигаться относительно окружающих ионов Ca2+. Новая молекулярная компонента будет иметь способность повышать нагрузку на мышцы и увеличивать их силу и выносливость.
Это улучшение могло бы предоставить дополнительные источники энергии на молекулярном уровне, повысить мощность сокращения мышцы и обеспечить более эффективное использование энергии. Более детальные исследования и эксперименты позволят более полно оценить потенциальные преимущества и возможные негативные последствия такой модификации молекулярного механизма сокращения мышцы.
Примечания:
- Литература, основанная на исследованиях:
- Грюнбаум Б., Емерисон Д., Джаррелл Д. Основы биохимии мышечных сокращений.
- Смит М., Куин Д. Молекулярная биология мышц.
- Траммел М. Механизмы сокращения и регулирования скорости сокращения скелетных мышц.
0 Комментариев